

Study on Activity Rhythm of Asiatic Black Bear in Shennongjia National Park Pilot Area
-
摘要:
神农架国家公园体制试点区(以下简称“神农架国家公园”)位于我国华中区域,是亚洲黑熊(Ursus thibetanus)的重要分布区域。目前,缺乏该物种在神农架林区的活动节律研究, 无法支撑神农架国家公园制定该物种针对性的保护方案。研究收集了2018—2020年神农架国家公园772台次红外相机位点监测数据,累计62 575个相机工作日,利用其中亚洲黑熊的影像数据分析其活动节律,并采用卡方检验和单因素方差分析法分析亚洲黑熊对不同海拔段的利用强度及不同季节的垂直迁移行为。结果表明: (1)神农架国家公园亚洲黑熊年活动节律为单峰型,夏季和秋季的活动强度较高,冬眠期为12月下旬至次年4月中旬,持续时间约为3个半月。(2)神农架林区亚洲黑熊日活动节律为晨昏活动模式,日间活动强度更高,具有明显的昼行性。(3)在海拔梯度上,亚洲黑熊在神龙架>1 000~2 600 m海拔段最为活跃。(4) 不同季节亚洲黑熊活动的海拔范围差异明显,具有垂直迁移行为。(5)在神农架林区亚洲黑熊拍摄率最低的>1 600~1 800 m海拔段内, 公路长度和居民点数量分别居第2和第1,说明人为活动对亚洲黑熊活动产生较大影响。神农架国家公园亚洲黑熊的活动节律研究具有重要价值,可为神农架国家公园制定亚洲黑熊种群的保护管理方案以及缓解人熊冲突提供重要的基础数据和科学支撑。
Abstract:Pilot area of Shennongjia National Park (SNP) contains important habitat of Asiatic black bear (Ursus thibetanus) in central China, however, there is a lack of research on the activity rhythm of this species in SNP, which is conducive for managers to make better management plan for conserving the species. Occurrence data of Asiatic black bear were collected from 772 camera trapping sites from 2018 to 2020 with 62 575 camera trapping nights. Date and time of photographs of Asiatic black bear were extracted to assess their activity rhythm. The chi-square test and one-way ANOVA test were used to analyze the behavior of black bear at different elevations and vertical migration in different seasons. Below are the research results: (1) The annual activity rhythm of Asiatic black bears in SNP is unimodal, with the highest activity intensity occurring in summer and autumn. The hibernation period of black bear lasts from late December to mid-April of the next year for about 3 and a half months. (2) Daily activity rhythm of Asiatic black bear in SNP was bimodal, and the intensity of diurnal activity was higher, showing obvious diurnal pattern. (3) Asiatic black bear of SNP was most active between the altitude from >1 000 to 2 600 m. (4) There were significant differences in the range of elevations for the activity of Asiatic black bear in SNP in different seasons, which showed obvious vertical migration behavior of the species. (5) The lowest photograph rate of Asiatic black bear in SNP occurred between the altitude of >1 600-1 800 m, as the length of highway and the number of settlements in this elevation section ranked the second and first in SNP, respectively, indicating that human activities had a great influence on Asiatic black bear activities. The findings on the activity rhythm of the Asiatic black bears will provide important basic data and scientific support for the conservation and management of Asiatic black bear population and mitigation of human-bear conflict in SNP.
-
Keywords:
- Asiatic black bear /
- camera trap /
- elevation /
- hibernation /
- activity rhythm /
- Shennongjia National Park
-
动物活动节律是动物行为生态学的一个重要内容,主要研究动物在不同时间内的活动强度及变化规律[1]。活动节律是动物对各种环境周期性变化的一种综合性适应,包括对光、温度、湿度等非生物条件和食物条件、种内社群关系和天敌等种间关系的适应[2],能反映动物个体的营养状态、社会地位、生存压力等参数[3]。食物的可获得性和丰度可以影响黑熊的季节性和日常活动模式[4-5],另外人为活动(如修建道路、栖息地变化等)也会对亚洲黑熊(Ursus thibetanus) 产生威胁,影响其日常的活动模式[6]。当前对于亚洲黑熊活动节律的研究主要集中于北温带分布区[4-5]、低纬度高海拔温带森林分布区[7]、北温带亚热带交界森林分布区[8]以及亚热带森林分布区[6],主要关注食性对亚洲黑熊活动节律的影响[5, 9-10]。而对于亚洲黑熊在中纬度地区唯一保存完好的原始森林生态区的活动节律研究较少,因而对该地区亚洲黑熊活动节律进行研究,分析该珍稀物种的生存对策和受威胁现状,可为制定减少人熊冲突的防护策略提供依据,促进其保护管理。
亚洲黑熊隶属于食肉目(Carnivora)熊科(Ursidae)熊属(Ursus),在地理分布上属于东洋界物种,为亚洲特有种[11],分布于四川盆地周缘山地和大巴山系的亚洲黑熊栖息于海拔1 000~2 600 m的中山常绿阔叶林和针阔混交林中[12]。亚洲黑熊分布于亚洲的17个国家和地区,从东亚至南亚北部及东南亚都有分布[13]。目前对活动节律的研究详细的亚种主要为分布于北温带的日本亚种(U. t. japonicus)[5, 14]、亚热带中国台湾的台湾亚种(U. t. formosanusbao)[4]、云南地区四川亚种(U. t. mupinensis) [7]和秦岭地区四川亚种[8]。亚洲黑熊在日本为全天活动节律,白天活动强度低于夜晚[5]; 亚洲黑熊在我国台湾地区活动存在季节差异,春夏季白天活动强度高于夜晚,秋季白天活动强度低于夜晚[4]。云南地区亚洲黑熊的晨昏活动无昼夜差异,且夏季活动更为活跃[7]。秦岭地区亚洲黑熊各季节活动呈现多个高峰, 秋季活动更为频繁[8]。亚洲黑熊四川亚种具有冬眠习性,夏秋季节大量进食,为冬眠储存脂肪,冬眠结束时间为次年3—4月[4]。目前,对于北亚热带季风气候区,尤其是亚热带气候向温带气候过渡区域森林亚洲黑熊的活动节律研究存在欠缺,不利于该物种的保护与管理。
神农架国家公园体制试点区(以下简称“神农架国家公园”)属大巴山系,地势西南高东北低,地貌类型复杂且生物多样性丰富,是亚洲黑熊的重要栖息地分布区[15-16]。神农架国家公园开展了一系列的生物多样性保护行动,但仍存在旅游、采集、偷猎等人为活动[17-18], 对野生动植物保护和管理具有一定的干扰和影响。为实现神农架地区人熊共存,亟需开展亚洲黑熊的生态学研究。因此,研究利用红外相机技术对神农架国家公园范围内的亚洲黑熊开展调查,研究其活动节律,分析影响其活动节律的因素,特别是人为活动对黑熊活动节律的影响,以期为该地区亚洲黑熊种群的保护以及缓解人熊冲突提供重要的理论依据和参考。
1. 材料与方法
1.1 研究区概况
神农架国家公园位于湖北省西北部神农架林区西南部地区(31°21′24.223″~31°36′27.317″ N,109°56′3.347″~110°36′26.779″ E),地处长江与汉水的分水岭,总面积1 170 km2,占神农架林区总面积的35.97%。神农架是我国北亚热带山地中在物种多样性、生态系统类型、生物生态学过程等方面具有国际意义的生物多样性保护的关键地区之一,也是具有地理标志意义的最重要的国家生态安全屏障区之一[19-20]。神农架国家公园垂直高差达2 700 m,神农顶海拔3 106.2 m,为大巴山脉主峰和湖北省的最高点[21]。神龙架受亚热带环流控制,属北亚热带季风气候区,温暖湿润,为中国南部亚热带与北部暖温带的气候过渡区域,从低海拔到高海拔依次呈现出北亚热带、暖温带、温带、寒温带的气候特点,形成了从低海拔到高海拔完整的山地植被垂直带系统[21-22]。
神农架国家公园内动物资源丰富,记录到哺乳纲动物89种、鸟纲动物443种,其中国家Ⅰ级重点保护动物9种,包括川金丝猴(Rhinopithecus roxellane)、豹(Panthera pardus)、东方白鹳(Ciconia boyciana)等; 国家Ⅱ级重点保护动物80种,包括亚洲黑熊、猕猴(Macaca mulatta)、藏酋猴(Macaca thibetana)、大灵猫(Viverra zibetha)等[23]。
1.2 布设相机
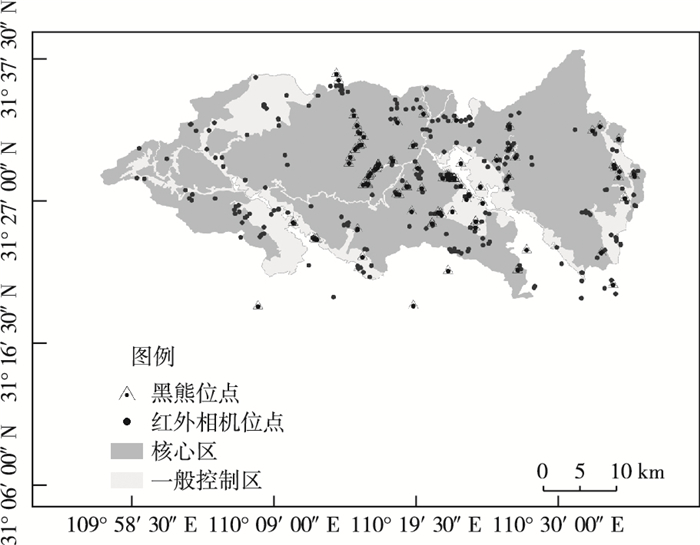
2017年10月—2020年10月,神农架国家公园的工作人员在大九湖管理区、神农顶管理区、木鱼管理区、老君山管理区4个片区内的18个管护中心管辖区域内共布设了178台红外相机。红外相机位置设置依据神农架国家公园范围内18个管护中心的巡护路线,每个管护分区的工作人员在所辖范围内不同海拔段以及典型植被类型中寻找野生动物活动痕迹比较多的区域布设红外相机,每次布设1台红外相机,每3个月定期检查维护回收1次红外相机数据,并及时清理红外相机前杂草等干扰物,每台相机轮换时间为6~9个月不等(图 1、表 1)。
![]() 图 1 神农架国家公园红外相机监测到亚洲黑熊的位点Figure 1. The locations of Asiatic black bears detected by camera traps in Shennongjia National Park system pilot area表 1 2018—2020年神农架国家公园红外相机工作日及获得的亚洲黑熊独立照片数Table 1. Number of camera trapping nights and Asiatic black bears independent photos captured by camera traps in Shennongjia National Park system pilot area from 2018 to 2020
图 1 神农架国家公园红外相机监测到亚洲黑熊的位点Figure 1. The locations of Asiatic black bears detected by camera traps in Shennongjia National Park system pilot area表 1 2018—2020年神农架国家公园红外相机工作日及获得的亚洲黑熊独立照片数Table 1. Number of camera trapping nights and Asiatic black bears independent photos captured by camera traps in Shennongjia National Park system pilot area from 2018 to 2020海拔/m 相机日/d 独立照片数 海拔/m 相机日/d 独立照片数 600 162 0 1 900 1 836 1 700 620 7 2 000 2 134 1 800 1 348 1 2 100 2 834 11 900 1 376 7 2 200 5 632 13 1 000 2 459 12 2 300 2 129 21 1 100 1 024 2 2 400 1 378 4 1 200 1 864 16 2 500 4 309 21 1 300 3 712 7 2 600 1 249 0 1 400 4 499 34 2 700 640 1 1 500 3 534 6 2 800 859 1 1 600 7 112 16 2 900 84 0 1 700 4 060 4 — 3 874 35 1 800 3 482 30 “—”表示红外相机中未记录海拔高度。 红外相机主要布设于野生动物出现可能性最大、动物活动痕迹明显且又能避开人类活动的地方。将红外相机绑到距离地面0.8~1.2 m高度的树上,树干直径大小合适,布设相机视角与兽径呈现一定俯角,且位点距离兽径、水源点或卧息点垂直距离2~5 m。红外相机安放的同时,由巡护人员在红外相机内和生境表中记录安放时间、相机编号、经纬度、海拔、森林植被类型等信息。相机参数设置为拍照+视频模式,灵敏度为中,每次感应触发连续拍摄3次加一段10 s视频。
1.3 数据处理与分析
对布设和回收数据与记录生境表信息对应,认真浏览每张照片并删除空照片,使用全国生物多样性监测平台红外相机数据处理工具(http://pbiomon.cn)对拍摄动物的照片进行物种识别鉴定,提取其中的亚洲黑熊信息数据。将同一相机位点含相同个体的相邻有效照片间隔时间30 min作为独立照片的确定标准,连续相邻照片时间≤30 min视为1个独立照片(independent photograph,IP)[24]。
采用动物的红外相机拍摄率(capture rate,CR, RC)作为表征相对多度的指标, 来分析某物种在不同时间的变化情况[25-27], 具体公式为
$$ R_{\mathrm{C}}=N_i \times 100 / T_i \times 100 \% 。 $$ (1) 式(1)中,Ni为该物种第i个月拍摄到的独立有效照片总数;Ti为第i个月的有效监测日。拍摄率越大,表明物种在该月份活动越频繁。
使用R软件的overlap包、activity包以及openxlsx包[28],以核密度估计法分析红外相机拍摄的独立有效照片的时间数据。首先使用R代码将独立有效照片的时间转化为弧度数据,并导入overlap包,使用densityPlot函数绘制单物种核密度曲线图,然后使用overlapEst函数计算重叠系数[28]。进一步分析亚洲黑熊的日活动节律,比较亚洲黑熊季节活动的日活动规律,实验步骤和R代码参考陈立军等[28]的研究。
根据研究区的气候特点,通过计算当地每日的日出和日落时间,统计每个季节的日出和日落平均时间(https://richurimo.bmcx.com/),神农架地区四季时间分别为春季3—5月、夏季6—8月、秋季9—11月、冬季12月—次年2月,不同季节的夜间时段划分标准为春季18:51—05:58、夏季19:18—05:34、秋季17:55—06:27、冬季17:46—07:12。利用夜间相对丰富度(night-time relative abundance index,NRAI, INRA)计算亚洲黑熊夜间活动时间占总日长的比例,进一步分析研究区内亚洲黑熊的夜行性[29],计算公式为
$$ I_{{\mathrm{NRA}}, i}=D_i / N_i \times 100 \%。 $$ (2) 式(2)中,i为季节,取值范围为0~3,0为平均值,1为春季,2为夏季,3为秋季(冬季亚洲黑熊具有冬眠行为);Di为某物种在i季节夜间时段的独立有效照片数;Ni为该物种i季节的独立有效照片总数。若某种动物NRAI值>13/24,则具有夜行性,反之则具有昼行性,接近则认为日节律没有昼夜差异。
采用Excel软件对记录到亚洲黑熊影像的红外相机生境坐标点对应的海拔进行数据处理,并利用SPSS 25.0软件中的非参数卡方检验分析亚洲黑熊对不同海拔段利用强度(独立照片数量比例)的差异,将神农架海拔按200 m的海拔间隔进行分段,海拔段以整数值表示,对比计算不同海拔段亚洲黑熊拍摄率,采用单因素方差分析法(one-way ANOVA)分析比较亚洲黑熊不同季节的海拔差异[7, 27]。
神农架国家公园30 m DEM高程数据由神农架国家公园管理局保护利用科提供,从全国地理信息资源目录服务系统(https://www.webmap.cn/main.do?method=index)获取2019年湖北省神农架地区1∶25万基础地理数据中的公路和居民点等信息。利用ArcGIS 10.7软件对神农架海拔段进行分段统计,利用IBM SPSS Statistics 25软件中的非参数检验分析公路长度和居民点数量对亚洲黑熊在不同海拔段活动强度的影响差异。
使用Arc GIS 10.7、Origin 2021、Adobe Illustrator 2021和R 3.6.3软件制图。
2. 结果与分析
2.1 亚洲黑熊年活动节律
此次红外相机调查研究历时3 a,共在神农架国家公园布设了772个位点的红外相机,累计62 575个相机工作日。其中共有93台次相机拍摄到亚洲黑熊照片,获得亚洲黑熊记录497条,相机内置时间错误数据4条,处理后获得亚洲黑熊独立照片251张。
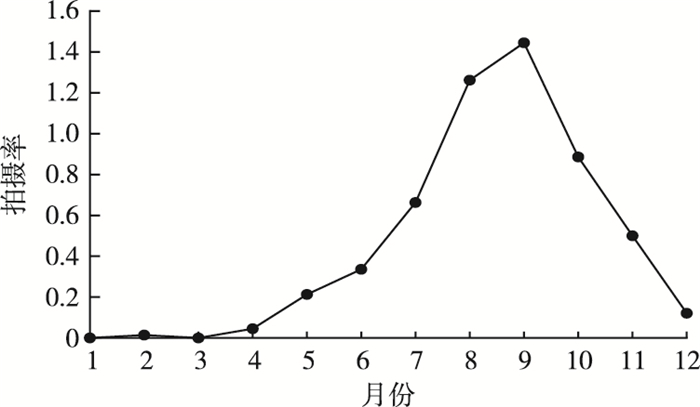
神农架国家公园亚洲黑熊的年活动强度变化结果(图 2)显示,亚洲黑熊在4月活动强度开始上升,6—7月活动急剧增加,8—9月处于活动高峰期,9月之后亚洲黑熊活动开始急剧降低,1—3月几乎无亚洲黑熊出现。2018—2020年仅在2月拍摄到1次亚洲黑熊活动记录。亚洲黑熊的主要活动时期为夏秋两季(7—11月),冬眠期为1—4月中旬,冬眠期时长3.5个月。
![]() 图 2 神农架国家公园亚洲黑熊的年活动节律Figure 2. Annual rhythm of Asiatic black bears in Shennongjia National Park system pilot area
图 2 神农架国家公园亚洲黑熊的年活动节律Figure 2. Annual rhythm of Asiatic black bears in Shennongjia National Park system pilot area2.2 亚洲黑熊日活动规律及季节性差异分析
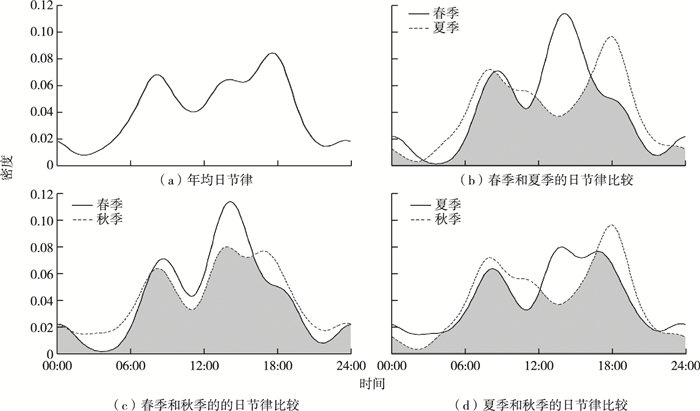
神农架国家公园亚洲黑熊日相对活动强度分析结果(图 3)显示,亚洲黑熊日活动为典型的晨昏活动模式。亚洲黑熊的日活动规律呈现双峰活动模式,高峰期出现在06:00—10:00和16:00—18:00,活动低谷出现在10:00—12:00和22:00—04:00;且日间活动强度更高,具有较为明显的昼行性,下午的活动密度峰期是上午的2倍左右。
![]() 图 3 神农架国家公园亚洲黑熊的活动节律Figure 3. Activity rhythm of Asiatic black bears in Shennongjia National Park system pilot area
图 3 神农架国家公园亚洲黑熊的活动节律Figure 3. Activity rhythm of Asiatic black bears in Shennongjia National Park system pilot area分析亚洲黑熊不同季节的日活动强度规律(图 3)可知,春季、夏季和秋季的日节律差异明显(P < 0.01),其中春季、夏季和秋季独立照片数量分别为14、94和97,夏季和秋季的活动强度高且活动差异明显。春季、夏季、秋季的NRAI值分别为0.143、0.154、0.250,说明亚洲黑熊的日活动具有明显的昼行性。
2.3 亚洲黑熊海拔利用偏好分析
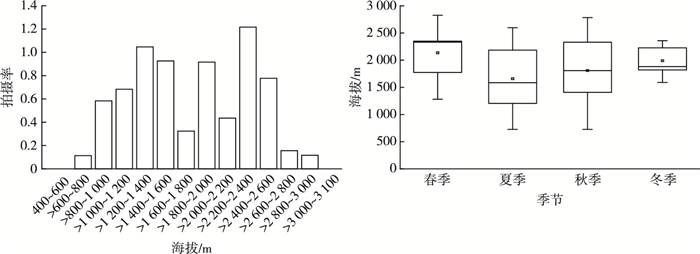
神农架亚洲黑熊对不同海拔的利用频率存在显著差异(χ2=149.68,P<0.01)。红外相机记录的亚洲黑熊活动范围为726~2 826 m,且主要集中在>1 000~2 600 m海拔范围内,其中亚洲黑熊在>1 200~1 600、>1 800~2 000、>2 200~2 600 m海拔段内拍摄率较高(图 4)。神农架国家公园亚洲黑熊在各季节的活动海拔均值分别为春季(2 330.0±478.46) m、夏季(1 658.28±505.29) m、秋季(1 808.30±581.43) m、冬季(1 990.91±270.54) m,亚洲黑熊在最活跃的夏季和秋季主要在海拔>1 000~2 600 m活动。
![]() 图 4 神农架国家公园亚洲黑熊的主要活动海拔和不同季节活动海拔分布上下线分别表示亚洲黑熊活动的最高和最低海拔,中间线表示亚洲黑熊活动海拔的中位数,数据点表示平均海拔。Figure 4. The main activity altitude of Asiatic black bears and the activity altitude of different seasons in Shennongjia National Park system pilot area
图 4 神农架国家公园亚洲黑熊的主要活动海拔和不同季节活动海拔分布上下线分别表示亚洲黑熊活动的最高和最低海拔,中间线表示亚洲黑熊活动海拔的中位数,数据点表示平均海拔。Figure 4. The main activity altitude of Asiatic black bears and the activity altitude of different seasons in Shennongjia National Park system pilot area亚洲黑熊在不同季节的活动海拔范围具有明显差异,春季活动海拔区间最低值1 281 m,最高值2 826 m,中位数2 330 m,IQR值575; 夏季活动海拔区间最低值726 m,最高值2 597 m,中位数1 585.6 m,IQR值979; 秋季活动海拔区间最低值726 m,最高值2 783 m,中位数1 808.9 m,IQR值962.5; 冬季活动海拔区间最低值1 589 m,最高值2 359 m,中位数1 883.2 m,IQR值408。说明亚洲黑熊具有垂直迁移行为,海拔差异显著(F值为3.428,df值为3,P值为0.018)。
2.4 公路和居民点对亚洲黑熊活动节律的影响
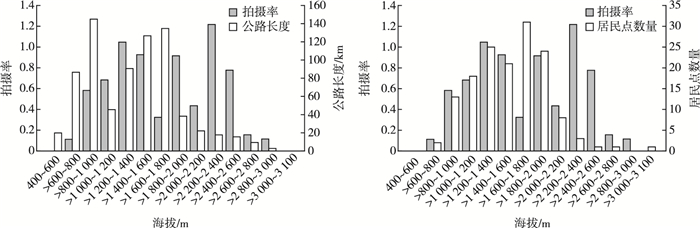
神农架国家公园内的公路长度(χ2=38.25,P=0.04)和居民点数量(χ2=8, P=0.21)对亚洲黑熊活动产生明显影响。根据神农架海拔段对公路长度和居民点数量进行划分,400~1 600 m海拔段公路长度和居民点占比较大,随海拔增加占比整体呈递增趋势,与亚洲黑熊拍摄率呈正相关。在>1 600~1 800 m海拔段,公路长度和居民点数量占比分别居第2和第1位,此海拔段的亚洲黑熊拍摄率最低。在>1 800~3 100 m海拔段,公路长度和居民点数量占比均较小,且随海拔增加,占比整体呈递减趋势,与亚洲黑熊拍摄率呈负相关(图 5)。
![]() 图 5 神农架国家公园不同海拔段内公路和居民点分布对拍摄率的影响Figure 5. Distribution of highway and residential Areas in Shennongjia National Park system pilot area
图 5 神农架国家公园不同海拔段内公路和居民点分布对拍摄率的影响Figure 5. Distribution of highway and residential Areas in Shennongjia National Park system pilot area3. 讨论
神农架国家公园亚洲黑熊的日活动类型具有典型的晨昏活动模式,日活动规律为双峰活动模式,其中黑熊的日活动高峰期出现在06:00—10:00和16:00—18:00,活动低谷出现在10:00—12:00和22:00—04:00; 具有较为明显的昼行性。这与唐家河保护区的四川亚种和日本亚种相似,其在黎明和黄昏活动明显[5, 30],且各季节的的日活动模式与陕西南部秦岭地区的四川亚种相似[8],但是与云南省西北部云岭自然保护区的四川亚种和中国台湾的台湾亚种略有不同[4, 7]。云岭自然保护区四川亚种的活动高峰期为06:00—09:00和20:00—23:00[7]; 我国台湾亚种活动高峰期为06:00—16:00,且06:00达活动强度最高值,24:00达活动强度最低值[4],这可能与其所分布区域的气候差异有关。
食物是影响亚洲黑熊活动节律的重要因素之一,黑熊是杂食性的机会主义捕食者,会随季节改变它们的栖息地和食物习惯[31-32]。亚洲黑熊因具有冬眠的习性,在春冬两季的日活动规律不明显,频繁活动的季节为夏季和秋季。春季(3—5月)亚洲黑熊主要以绿色植物和群居小动物为食,如多汁的草本植物、前一年的橡子和蚂蚁等,但也有饮食行为研究发现熊在春天有捕食鹿的行为[9, 33-34]。夏季(6—9月),亚洲黑熊初期以富含糖的浆果、多汁嫩草、蜂蜜等为食,后期以玉米、土豆、树果、藤本果实等为食[33, 35-37]。秋季(9—11月)亚洲黑熊以橡子、板栗等坚果为主要食物,为冬眠储存脂肪[9, 36, 38]。春季神农架地区亚洲黑熊结束冬眠后开始外出进行活动,嫩叶、竹笋等亚洲黑熊的食物开始出现,亚洲黑熊外出活动开始增加且主要为白天活动,觅食活动高峰为13:00—16:00。6月左右为神农架地区蜜蜂产蜜期,多处出现亚洲黑熊肇事搬食蜂蜜事件,之后林内和农作物区内开始出现亚洲黑熊啃食玉米、土豆、五味子、竹笋、树果等觅食痕迹。亚洲黑熊的主要食源——锐齿槲栎、橡子树和野生核桃等坚果树木以及野生猕猴桃等野果进入成熟期后,亚洲黑熊开始大量进食,处于活动高峰期。11—12月亚洲黑熊活动强度下降,12月底左右进入冬眠期。
印度亚洲黑熊指名亚种(U. t. thibetanus)(活动海拔1 500~4 610 m)9—10月也会根据橡子采食地点的变化从高海拔向低海拔进行垂直迁移[32]。印度支那地区老挝南富路易国家保护区亚洲黑熊指名亚种(活动海拔400~2 257 m)雨季的活动密度会比低海拔地区高[39]。伊朗亚洲黑熊喜马拉雅亚种(U. t. laniger)(活动海拔428~1 878 m)在霍尔木兹甘省的季节性觅食地椰枣林成熟时,会从高海拔林地向河岸地区迁移觅食[40]。而伊朗地区巴基斯坦亚种(U. t. gedrosianus)因气候变化和栖息地破坏等原因,会向北部和西部海拔较高的地区迁移,适宜生境的海拔高度范围从目前的200~2 000 m向未来的200~2 500 m转变[41]。日本亚洲黑熊日本亚种(活动海拔600~3 000 m)因食源地点的变化会从高海拔迁移至低海拔地区,存在明显的垂直迁移[36, 42-43]。针对中国东北(活动海拔115~821 m)和俄罗斯的亚洲黑熊东北亚种(U. t. ussuricus)的研究记录到其因栖息地变化而进行的水平迁移[44-45],与尼泊尔(活动海拔1 900~3 100 m)和巴基斯坦(活动海拔2 500~3 000 m)分布的亚洲黑熊指名亚种一样,暂时没有记录到食源地季节性变化引起的垂直迁移[46-47]。经过对比研究发现,活动海拔跨度越大越促使亚洲黑熊进行海拔垂直迁移,同时独特的气候变化、植被垂直带和地形条件等综合因素对亚洲黑熊的垂直迁移行为产生影响。神农架国家公园亚洲黑熊在不同季节的活动海拔范围具有明显差异。神农架国家公园森林覆盖率高,因其独特的气候变化和地理位置拥有从低海拔到高海拔完整的山地植被垂直带系统,这可能是亚洲黑熊具有明显的垂直迁移行为的驱动力,这一现象与已有研究观测到的云南亚洲黑熊四川亚种(活动海拔2 900~3 800 m)、四川亚洲黑熊四川亚种(活动海拔1 500~2 500 m)没有垂直迁移行为报道不一致[7, 35]。
野生动物活动节律既是动物本身进化适应的一部分,又是其对环境因素变化采取生存对策的一种适应[48]。亚洲黑熊的活动节律不可避免会受到神农架国家公园内人为活动的影响和干扰。神农架国家公园范围内有高等级生态旅游专线公路、依托旅游景点发展的旅游小镇和居民点。在亚洲黑熊主要活动海拔带中,>1 000~1 600 m范围内公路长度和居民点数量占比较大,但该海拔段内的常绿落叶阔叶混交林为亚洲黑熊的食源地,故亚洲黑熊依旧保持较高的活动强度。但是在>1 600~1 800 m范围内公路长度和居民点数量占比较高,而此海拔段的亚洲黑熊拍摄率最低,表明人为活动对亚洲黑熊活动产生较大影响。尤其是在旅游高峰期人流量和车流量较大,人为活动干扰不可避免对亚洲黑熊的活动带来影响,这可能也是亚洲黑熊具有明显的垂直迁移行为的驱动力之一。
神农架国家公园属北亚热带季风气候区(31°21′20″~31°36′20″ N),神农架亚洲黑熊从每年4月中旬开始活动增强,表明冬眠结束时间在4月(2月2日出现的个例除外),最晚出现时间为12月底,推测亚洲黑熊冬眠期约3.5个月。亚洲黑熊的冬眠存在纬度差异,亚洲黑熊分布的纬度越高,其冬眠时间开始越早,且持续时间越长[6]。在以往的亚洲黑熊越冬研究中, 相近纬度的四川卧龙自然保护区(30°45′~ 31°25′ N)和唐家河自然保护区(32°32′~32°41′ N)分布的亚洲黑熊冬眠期约为11月下旬至4月上旬[35];在陕西秦岭南坡相邻的观音山国家级自然保护区、佛坪国家级自然保护区、长青国家级自然保护区(33°19′~33°46′ N)分布的亚洲黑熊冬眠期约为12月中旬开始,1—3月活动较少,且秦岭地区的气候条件比热带和亚热带地区更冷,其冬眠期至少在3个月以上[8]。神农架亚洲黑熊冬眠时间和模式与研究记录的亚洲黑熊冬眠纬度模式相吻合。神农架国家公园立体地形小气候明显,且随着海拔逐渐增高,10月至次年4月期间神农架地区依次迭现暖温带、中温带、寒潮带等多种气候类型。10月之后神农架地区温度逐渐降低,食物和光照减少,亚洲黑熊逐渐进入冬眠期。次年3—5月(春季期间)亚洲黑熊结束冬眠,开始外出进行活动,但因为神农架地理位置特殊,海拔高度差异较大,且3—5月温度变化较大,天气多变(4月底神农顶等高海拔地区还会出现降雪现象),影响亚洲黑熊的外出活动,但随着温度升高,嫩叶、竹笋等亚洲黑熊的食物开始出现,促使亚洲黑熊外出活动频率开始增加。
亚洲黑熊活动节律是亚洲黑熊对环境适应的表现,对亚洲黑熊活动节律的研究有利于神农架国家公园有针对性地制定亚洲黑熊的保护方案。监测分析亚洲黑熊的活动节律与活动范围,利于亚洲黑熊的控制与管理。在神农架国家公园内的实际调查访问表明,人兽冲突事件中亚洲黑熊作为肇事野生动物,以破坏蜂农的蜂箱取食蜂蜜和破坏庄稼取食玉米为主。在夏秋两季亚洲黑熊的活动高峰加强监测,有利于规避亚洲黑熊伤人的风险。在亚洲黑熊活动频繁区域的蜂场修建高架平台保护蜂箱,种植地可选择种植茶叶或者药材等高经济作物,以减轻亚洲黑熊对农作物的破坏。神农架国家公园内亚洲黑熊的日活动高峰期出现在06:00—10:00和16:00—18:00,这段时间主要为亚洲黑熊的觅食期,建议进一步控制神农架国家公园对游客开放时间、当地居民进山劳作和采集等人为活动时间,规避野生动物的活动高峰期,以利于减少人为活动对亚洲黑熊等野生动物活动节律的干扰。因此,建议在之后的工作中持续对野生动物尤其是亚洲黑熊进行系统监测,对于野外红外相机监测体系采用科学的网格化安放以建立长期监测固定样方,为评估长期趋势定期开展兽类调查,以便更深入地研究亚洲黑熊的种群、行为、活动节律等特征。
致谢: 野外调查得到湖北省神农架国家公园管理局的大力支持,感谢神农架国家公园的工作人员在野外调查工作中的辛苦付出。 -
![]()
图 1 神农架国家公园红外相机监测到亚洲黑熊的位点
Figure 1. The locations of Asiatic black bears detected by camera traps in Shennongjia National Park system pilot area
![]()
图 2 神农架国家公园亚洲黑熊的年活动节律
Figure 2. Annual rhythm of Asiatic black bears in Shennongjia National Park system pilot area
![]()
图 3 神农架国家公园亚洲黑熊的活动节律
Figure 3. Activity rhythm of Asiatic black bears in Shennongjia National Park system pilot area
![]()
图 4 神农架国家公园亚洲黑熊的主要活动海拔和不同季节活动海拔分布
上下线分别表示亚洲黑熊活动的最高和最低海拔,中间线表示亚洲黑熊活动海拔的中位数,数据点表示平均海拔。
Figure 4. The main activity altitude of Asiatic black bears and the activity altitude of different seasons in Shennongjia National Park system pilot area
![]()
图 5 神农架国家公园不同海拔段内公路和居民点分布对拍摄率的影响
Figure 5. Distribution of highway and residential Areas in Shennongjia National Park system pilot area
表 1 2018—2020年神农架国家公园红外相机工作日及获得的亚洲黑熊独立照片数
Table 1 Number of camera trapping nights and Asiatic black bears independent photos captured by camera traps in Shennongjia National Park system pilot area from 2018 to 2020
海拔/m 相机日/d 独立照片数 海拔/m 相机日/d 独立照片数 600 162 0 1 900 1 836 1 700 620 7 2 000 2 134 1 800 1 348 1 2 100 2 834 11 900 1 376 7 2 200 5 632 13 1 000 2 459 12 2 300 2 129 21 1 100 1 024 2 2 400 1 378 4 1 200 1 864 16 2 500 4 309 21 1 300 3 712 7 2 600 1 249 0 1 400 4 499 34 2 700 640 1 1 500 3 534 6 2 800 859 1 1 600 7 112 16 2 900 84 0 1 700 4 060 4 — 3 874 35 1 800 3 482 30 “—”表示红外相机中未记录海拔高度。  下载: 导出CSV
下载: 导出CSV
-
[1] 原宝东, 孔繁繁. 哺乳动物活动节律研究进展[J]. 安徽农业科学, 2011, 39(2): 1056-1058, 1162. DOI: 10.13989/j.cnki.0517-6611.2011.02.025 YUAN Bao-dong, KONG Fan-fan. Research on Mammal Activity Rhythm[J]. Journal of Anhui Agricultural Sciences, 2011, 39(2): 1056-1058, 1162. DOI: 10.13989/j.cnki.0517-6611.2011.02.025
[2] 孙儒泳, 王德华, 牛翠娟. 动物生态学原理[M]. 4版. 北京: 北京师范大学出版社, 2019: 21-224. [3] 陈梦霄, 王晓玲, 穆丽光, 等. 内蒙古赛罕乌拉国家级自然保护区狍的日活动节律及其季节变化研究[J]. 四川动物, 2019, 38(2): 214-219. https://www.cnki.com.cn/Article/CJFDTOTAL-SCDW201902019.htm CHEN Meng-xiao, WANG Xiao-ling, MU Li-guang, et al. Daily and Seasonal Activity Rhythm of Capreolus Pygargus in Inner Mongolia Saihanwula National Nature Reserve[J]. Sichuan Journal of Zoology, 2019, 38(2): 214-219. https://www.cnki.com.cn/Article/CJFDTOTAL-SCDW201902019.htm
[4] HWANG M H, GARSHELIS D L. Activity patterns of Asiatic black bears (Ursus thibetanus) in the Central Mountains of Taiwan[J]. Journal of Zoology, 2007, 271(2): 203-209. DOI: 10.1111/j.1469-7998.2006.00203.x
[5] YAMAZAKI K, KOZAKAI C, KASAI S, et al. A Preliminary Evaluation of Activity-sensing GPS Collars for Estimating Daily Activity Patterns of Japanese Black Bears[J]. Ursus, 2008, 19(2): 154-161. DOI: 10.2192/07GR009.1
[6] LIU F, MCSHEA W, GARSHELIS D, et al. Spatial Distribution as a Measure of Conservation Needs: An Example with Asiatic Black Bears in South-western China[J]. Diversity and Distributions, 2009, 15(4): 649-659. DOI: 10.1111/j.1472-4642.2009.00571.x
[7] 张永俊, 和育超, 赵娟钧, 等. 云岭自然保护区拉沙山区域亚洲黑熊的活动模式[J]. 兽类学报, 2021, 41(2): 136-143. https://www.cnki.com.cn/Article/CJFDTOTAL-SLXX202102003.htm ZHANG Yong-jun, HE Yu-chao, ZHAO Juan-jun, et al. Activity Patterns of Asiatic Black Bear (Ursus thibetanus) on the Mt. Lasha Yunling Nature Reserve[J]. Acta Theriologica Sinica, 2021, 41(2): 136-143. https://www.cnki.com.cn/Article/CJFDTOTAL-SLXX202102003.htm
[8] ZAHOOR B, LIU X H, WU P F, et al. Activity Pattern Study of Asiatic Black Bear (Ursus thibetanus) in the Qinling Mountains, China, by Using Infrared Camera Traps[J]. Environmental Science and Pollution Research, 2021, 28(20): 25179-25186. DOI: 10.1007/s11356-020-12325-3
[9] HASHIMOTO Y. Seasonal Food Habits of the Asiatic Black Bear (Ursus thibetanus) in the Chichibu Mountains, Japan[J]. Mammal Study, 2002, 27(1): 65-72. DOI: 10.3106/mammalstudy.27.65
[10] MORI T, SUGIURA R, KATO M, et al. A Seven-Year Longitudinal Study on the Food Habits of the Asiatic Black Bear (Ursus thibetanus) in Relation to Mast Production in Shirakawa Village, Gifu Prefecture, Japan[J]. Mammal Study, 2018, 43(2): 81-90. DOI: 10.3106/ms2017-0034
[11] 侯万儒, 胡锦矗. 中国熊类资源及其保护现状[J]. 四川师范学院学报(自然科学版), 1997, 18(4): 287-291. https://www.cnki.com.cn/Article/CJFDTOTAL-IGNE704.002.htm HOU Wan-ru, HU Jin-chu. The Present Situations of China's Bear Resources and Protection[J]. Journal of Sichuan Teachers College (Natural Science), 1997, 18(4): 287-291. https://www.cnki.com.cn/Article/CJFDTOTAL-IGNE704.002.htm
[12] 鲁庆彬, 胡锦矗. 岷山黑熊生境选择的初步分析[J]. 兽类学报, 2003, 23(2): 98-103. https://www.cnki.com.cn/Article/CJFDTOTAL-SLXX200302001.htm LU Qing-bin, HU Jin-chu. Preliminary Analysis on the Habitat Selection of Black Bears in the Minshan Mountains[J]. Acta Theriologica Sinica, 2003, 23(2): 98-103. https://www.cnki.com.cn/Article/CJFDTOTAL-SLXX200302001.htm
[13] GARSHELIS D, STEINMETZ R. Ursus thibetanus (Amended Version of 2016 Assessment)[EB/OL]. [2022-01-02]. https://dx.doi.org/10.2305/IUCN.UK.2020-3.RLTS.T22824A166528664.en.
[14] MCLELLAN M L, MCLELLAN B N. Effect of Season and High Ambient Temperature on Activity Levels and Patterns of Grizzly Bears (Ursus arctos)[J]. PLoS One, 2015, 10(2): e0117734. DOI: 10.1371/journal.pone.0117734
[15] 张如松, 杨其仁, 何定富, 等. 湖北省黑熊的种群动态[J]. 华中师范大学学报(自然科学版), 1995, 29(3): 378-381. https://www.cnki.com.cn/Article/CJFDTOTAL-HZSZ503.024.htm ZHANG Ru-song, YANG Qi-ren, HE Ding-fu, et al. Population Dynamics on Black Bear Resouces in Hubei Province[J]. Journal of Central China Normal University (Natural Sciences), 1995, 29(3): 378-381. https://www.cnki.com.cn/Article/CJFDTOTAL-HZSZ503.024.htm
[16] 戢太贵, 蒋国忠. 湖北神农架国家地质公园[J]. 资源环境与工程, 2006, 20(2): 207-208, 211. https://www.cnki.com.cn/Article/CJFDTOTAL-HBDK200602026.htm JI Tai-gui, JIANG Guo-zhong. Hubei Shennongjia National Geopark[J]. Resources Environment & Engineering, 2006, 20(2): 207-208, 211. https://www.cnki.com.cn/Article/CJFDTOTAL-HBDK200602026.htm
[17] 杨敬元, 廖明尧, 余辉亮, 等. 神农架金丝猴保护与研究现状[J]. 世界科技研究与发展, 2008, 30(4): 418-421. https://www.cnki.com.cn/Article/CJFDTOTAL-SJKF200804010.htm YANG Jing-yuan, LIAO Ming-yao, YU Hui-liang, et al. Protection and Research Status of Golden Monkeys in Shennongjia[J]. World SCI-TECH R & D, 2008, 30(4): 418-421. https://www.cnki.com.cn/Article/CJFDTOTAL-SJKF200804010.htm
[18] 张宇, 李丽, 张于光, 等. 人为干扰对神农架川金丝猴连通性及遗传多样性的影响[J]. 生态学报, 2019, 39(8): 2935-2945. https://www.cnki.com.cn/Article/CJFDTOTAL-STXB201908029.htm ZHANG Yu, LI Li, ZHANG Yu-guang, et al. Study on the Effect of Human Disturbance on the Connectivity and Genetic Diversity of Sichuan Snub-nosed Monkey (Rhinopithecus roxellana) in Shennongjia National Park[J]. Acta Ecologica Sinica, 2019, 39(8): 2935-2945. https://www.cnki.com.cn/Article/CJFDTOTAL-STXB201908029.htm
[19] 徐文婷, 谢宗强, 申国珍, 等. 神农架自然地域范围的界定及其属性[J]. 国土与自然资源研究, 2019(3): 42-46. https://www.cnki.com.cn/Article/CJFDTOTAL-GTZY201903011.htm XU Wen-ting, XIE Zong-qiang, SHEN Guo-zhen, et al. The Nature and Geography Scope of Shennongjia[J]. Territory & Natural Resources Study, 2019(3): 42-46. https://www.cnki.com.cn/Article/CJFDTOTAL-GTZY201903011.htm
[20] 谢宗强, 申国珍. 神农架国家公园体制试点特色与建议[J]. 生物多样性, 2021, 29(3): 312-314. https://www.cnki.com.cn/Article/CJFDTOTAL-SWDY202103012.htm XIE Zong-qiang, SHEN Guo-zhen. Distinguishing Feature and Suggestions of Shennongjia National Park System Pilot[J]. Biodiversity Science, 2021, 29(3): 312-314. https://www.cnki.com.cn/Article/CJFDTOTAL-SWDY202103012.htm
[21] 余小林, 韩文斌, 周友兵, 等. 神农架世界自然遗产地川金丝猴通道设计[J]. 生态科学, 2018, 37(4): 97-104. https://www.cnki.com.cn/Article/CJFDTOTAL-STKX201804012.htm YU Xiao-lin, HAN Wen-bin, ZHOU You-bing, et al. Technical Parameters for Designing Crossings for the Sichuan Snub-nosed Monkey(Rhinopithecus roxellana) in Shennongjia World Natural Heritage Site[J]. Ecological Science, 2018, 37(4): 97-104. https://www.cnki.com.cn/Article/CJFDTOTAL-STKX201804012.htm
[22] 马明哲, 申国珍, 熊高明, 等. 神农架自然遗产地植被垂直带谱的特点和代表性[J]. 植物生态学报, 2017, 41(11): 1127-1139. https://www.cnki.com.cn/Article/CJFDTOTAL-ZWSB201711001.htm MA Ming-zhe, SHEN Guo-zhen, XIONG Gao-ming, et al. Characteristic and Representativeness of the Vertical Vegetation Zonation along the Altitudinal Gradient in Shennongjia Natural Heritage[J]. Chinese Journal of Plant Ecology, 2017, 41(11): 1127-1139. https://www.cnki.com.cn/Article/CJFDTOTAL-ZWSB201711001.htm
[23] 周友兵, 韩文斌, 陈文文, 等. 神农架世界自然遗产地陆生脊椎动物多样性[J]. 生态科学, 2018, 37(5): 47-52. https://www.cnki.com.cn/Article/CJFDTOTAL-STKX201805007.htm ZHOU You-bing, HAN Wen-bin, CHEN Wen-wen, et al. Terrestrial Vertebrate Diversity in Shennongjia World Natural Heritage Site, China[J]. Ecological Science, 2018, 37(5): 47-52. https://www.cnki.com.cn/Article/CJFDTOTAL-STKX201805007.htm
[24] PALMER M S, SWANSON A, KOSMALA M, et al. Evaluating Relative Abundance Indices for Terrestrial Herbivores from Large-scale Camera Trap Surveys[J]. African Journal of Ecology, 2018, 56(4): 791-803.
[25] ROVERO F, MARTIN E, ROSA M, et al. Estimating Species Richness and Modelling Habitat Preferences of Tropical Forest Mammals from Camera Trap Data[J]. PLoS One, 2014, 9(7): e103300.
[26] 刘芳, 宿秀江, 李迪强, 等. 利用红外相机调查湖南高望界国家级自然保护区鸟兽多样性[J]. 生物多样性, 2014, 22(6): 779-784. https://www.cnki.com.cn/Article/CJFDTOTAL-SWDY201406016.htm LIU Fang, SU Xiu-jiang, LI Di-qiang, et al. Using Camera Trap to Investigate Animal Diversity in Hunan Gaowangjie National Nature Reserve[J]. Biodiversity Science, 2014, 22(6): 779-784. https://www.cnki.com.cn/Article/CJFDTOTAL-SWDY201406016.htm
[27] 李佳, 刘芳, 李迪强, 等. 基于红外相机监测分析的红腹角雉日活动节律[J]. 林业科学, 2017, 53(7): 170-176. https://www.cnki.com.cn/Article/CJFDTOTAL-LYKE201707017.htm LI Jia, LIU Fang, LI Di-qiang, et al. Daily Activity Rhythm of Temminick's Tragopan (Trgopan temminckii) Based on Infrared Camera Monitoring[J]. Scientia Silvae Sinicae, 2017, 53(7): 170-176. https://www.cnki.com.cn/Article/CJFDTOTAL-LYKE201707017.htm
[28] 陈立军, 束祖飞, 肖治术. 应用红外相机数据研究动物活动节律: 以广东车八岭保护区鸡形目鸟类为例[J]. 生物多样性, 2019, 27(3): 266-272. https://www.cnki.com.cn/Article/CJFDTOTAL-SWDY201903006.htm CHEN Li-jun, SHU Zu-fei, XIAO Hu-xiao. Application of Camera-trapping Data to Study Daily Activity Patterns of Galliformes in Guangdong Chebaling National Nature Reserve[J]. Biodiversity Science, 2019, 27(3): 266-272. https://www.cnki.com.cn/Article/CJFDTOTAL-SWDY201903006.htm
[29] 武鹏峰, 刘雪华, 蔡琼, 等. 红外相机技术在陕西观音山自然保护区兽类监测研究中的应用[J]. 兽类学报, 2012, 32(1): 67-71. https://www.cnki.com.cn/Article/CJFDTOTAL-SLXX201201009.htm WU Peng-feng, LIU Xue-hua, CAI Qiong, et al. The Application of Infrared Camera in Mammal Research in Guanyinshan Nature Reserve, Shaanxi[J]. Acta Theriologica Sinica, 2012, 32(1): 67-71. https://www.cnki.com.cn/Article/CJFDTOTAL-SLXX201201009.htm
[30] 胡锦矗, SCHALLER G B, JOHNSON K G. 唐家河自然保护区大熊猫的觅食生态研究[J]. 四川师范学院学报(自然科学版), 1990, 11(1): 1-13. https://www.cnki.com.cn/Article/CJFDTOTAL-IGNE199001000.htm HU Jin-chu, SCHALLER G B, JOHNSON K G. The Feeding Ecology of Giant Pandas in Tangjiahe Natural Reserve[J]. Journa of China West Normal University (Natural Sciences), 1990, 11(1): 1-13. https://www.cnki.com.cn/Article/CJFDTOTAL-IGNE199001000.htm
[31] HASHIMOTO Y, ANRUI T. Summer Food Habits of the Asiatic Black Bears in Moor of Oze Area, Central Japan[J]. Mammal Study, 2013, 38(4): 235-241.
[32] BASNETT R, KUMAR A, VISHWAKARM A A, et al. Seasonal Diets of Asiatic Black Bear (Ursus thibetanus) in the Khangchendzonga National Park, Eastern Himalaya India[J]. Journal of Natural History, 2021, 55(3/4): 163-175.
[33] KOIKE S, NAKASHITA R, KOZAKAI C, et al. Baseline Characterization of the Diet and Stable Isotope Signatures of Bears that Consume Natural Foods in Central Japan[J]. European Journal of Wildlife Research, 2016, 62(1): 23-31.
[34] FURUSAKA S, KOZAKAI C, NEMOTO Y, et al. The Selection by the Asiatic Black Bear (Ursus thibetanus) of Spring Plant Food Items According to Their Nutritional Values[J]. Zoo Keys, 2017, 672: 121-133.
[35] REID D, JIANG M, TENG Q, et al. Ecology of the Asiatic Black Bear (Ursus thibetanus) in Sichuan, China[J]. Mammalia, 1991, 55(2): 221-238.
[36] NAKAJIMA A, KOIKE S, MASAKI T, et al. Spatial and Elevational Variation in Fruiting Phenology of a Deciduous Oak (Quercus crispula) and Its Effect on Foraging Behavior of the Asiatic Black Bear (Ursus thibetanus)[J]. Ecological Research, 2012, 27(3): 529-538.
[37] MALCOLM K D, MCSHEA W J, GARSHELIS D L, et al. Increased Stress in Asiatic Black Bears Relates to Food Limitation, Crop Raiding, and Foraging beyond Nature Reserve Boundaries in China[J]. Global Ecology and Conservation, 2014, 2: 267-276.
[38] KOIKE S. Fruiting Phenology and Its Effect on Fruit Feeding Behavior of Asiatic Black Bears[J]. Mammal Study, 2009, 34(1): 47-52.
[39] SCOTSON L. The Distribution and Status of Asiatic Black bear Ursus thibetanus and Malayan Sun Bear Helarctos malayanus in Nam et Phou Louey National Protected Area, Lao PDR[R]. Minnesota, USA: Unpublished Report to International Association for Bear Research and Management, 2010.
[40] GHADIRIAN T, QASHQAEI A T, SOOFI M, et al. Diet of Asiatic Black Bear in Its Westernmost Distribution Range, Southern Iran[J]. Ursus, 2017, 28(1): 15-19.
[41] AZITA F. Modeling of Habitat Suitability of Asiatic Black Bear (Ursus thibetanus gedrosianus) in Iran in Future[J]. Acta Ecologica Sinica, 2018, 38(1): 9-14.
[42] HUYGENSOSCAR C, TOSHIRO M, BJØRN D, et al. Diet and Feeding Habits of Asiatic Black Bears in the Northern Japanese Alps[J]. Ursus, 2003, 14(2): 236-245.
[43] IZUMIYAMA S, SHIRAISHI T. Seasonal Changes in Elevation and Habitat Use of the Asiatic Black Bear (Ursus thibetanus) in the Northern Japan Alps[J]. Mammal Study, 2004, 29(1): 1-8.
[44] SKRIPOVA K V. Trends in the Spatial Distribution of Asiatic Black Bear Cubs[J]. Russian Journal of Ecology, 2009, 40(1): 61-64.
[45] 战如亮, 王文, 陈洋, 等. 黑龙江省通河县乌拉浑林场黑熊的生境选择[J]. 野生动物, 2010, 31(2): 69-73. https://www.cnki.com.cn/Article/CJFDTOTAL-YSDW201002005.htm ZHAN Ru-liang, WANG Wen, CHEN Yang, et al. Habitat Selection of Black Bears in Ulahun Forest Farm, Heilongjiang[J]. Chinese Journal of Wildlife, 2010, 31(2): 69-73. https://www.cnki.com.cn/Article/CJFDTOTAL-YSDW201002005.htm
[46] BISTA R, ARYAL A. Status of the Asiatic Black Bear Ursus thibetanus in the Southeastern Region of the Annapurna Conservation Area, Nepal[J]. Zoology and Ecology, 2013, 23(1): 83-87.
[47] ALI A, ZHOUZ X, WASEEM M, et al. An Assessment of Food Habits and Altitudinal Distribution of the Asiatic Black Bear (Ursus thibetanus) in the Western Himalayas, Pakistan[J]. Journal of Natural History, 2017, 51(11/12): 689-701.
[48] 易国栋, 杨志杰, 刘宇, 等. 中华秋沙鸭越冬行为时间分配及日活动节律. 生态学报, 2010, 30(8): 2228-2234. https://www.cnki.com.cn/Article/CJFDTOTAL-STXB201008034.htm YI G D, YANG Z J, LIU Y, et al. Behavioral Time Budget and Daily Rhythms for Wintering Mergus squamatus[J]. Acta Ecologica Sinica, 2010, 30(8): 2228-2234. https://www.cnki.com.cn/Article/CJFDTOTAL-STXB201008034.htm
-
期刊类型引用(2)
1. 杨柳,陈曦,王晓敏,向丰元,杨志斌,任泽梅. 神农架漆树资源调查及评价. 中国生漆. 2024(03): 29-33+48 .  百度学术
百度学术
2. 张留栓,刘芳,姬云瑞,陈静,李迪强,余辉亮,马国飞,杨敬元. 基于MaxEnt模型的神农架国家公园体制试点区的亚洲黑熊适宜生境研究. 陆地生态系统与保护学报. 2024(05): 14-24 . 百度学术
其他类型引用(1)
计量
- 文章访问数: 137
- HTML全文浏览量: 3
- PDF下载量: 24
- 被引次数: 3
 苏公网安备32000099338-24005号
苏公网安备32000099338-24005号